Microbes: tiny but mighty
Invisible to our eyes, microbes maintain energy flow in marine, freshwater, and inland sea food webs. Microbial communities are composed of diverse groups of Bacteria, Algae, and Fungi. In just a few places around the globe, microbial communities can build incredible structures that can be seen without the aid of a microscope!
Going back in time to understand the present
Microbes evolved billions of years ago, when Earth’s atmosphere was not hospitable to more complex life forms. Microbes living at that time had to figure out a way to survive in the face of adverse conditions. What are microbes to do during such dire times? Stick together! Microbes can stick together in cohesive mats, or biofilms, that allow the microbes to control the local environmental conditions. Maybe at one point you have seen green slimy stuff on the sides of a fish bowl; several microbes can secrete sticky glue called exopolymeric substances (EPS) that allows them to live as a biofilm attached to a surface. This glue is also important for trapping and binding minerals to build a more stable habitat for the microbial community, a process which over time can result in impressive stromatolite structures (Bowlin et al. 2012). Amazingly, this process of sticking together, which occurred eons ago, is still happening today!

Because microbes grew during adverse times, present-day microbes tend to thrive in “extreme” environments, such as hypersaline lakes and thermal springs, as well as marine systems (Fernandez et al. 2016). In a few extreme locations throughout the world, scientists have discovered remarkable types of biofilm communities, called microbialites.
Microbialites are reef-like structures that contain abundant communities of Algae, Bacteria, Fungi, and Archaea. Some of the most abundant microbialites can be found in marine coastal systems as well as “inland seas” such as Utah’s Great Salt Lake. Microbialites are increasingly regarded as important study systems to help us understand how microbes contribute to globally important processes such as carbon cycling, and they are also gaining attention as an important food source for aquatic organisms.
How do microbialites form?
The formation of microbialites depends on the interplay between the surrounding water chemistry and types of microbes floating around. Scientists often find microbialites in shallow, clear lakes and marine ecosystems that have many dissolved cations, such as calcium, as these are necessary for the reef-like part of microbialites. In addition, lakes and coastal areas with substantial groundwater input may have high numbers of cations and organic matter to fuel microbialite formation (Dupraz et al. 2009).
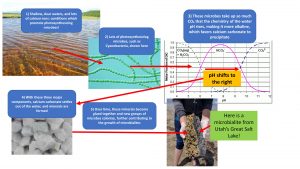
Cyanobacteria, a major group of bacteria, are dominant photoautotrophs (photosynthesizing organisms) that create localized increases in pH of the water. This happens because photosynthesis by Cyanobacteria consumes carbon dioxide, CO2, often at a faster rate than CO2 can be replaced from the atmosphere. This rapid depletion of CO2 changes the surrounding water chemistry and allows free-floating calcium to bond with carbonate molecules, becoming a solid mineral that falls out of the water column. This solid mineral is now ready to be colonized by microbes, and over time, more minerals form, which eventually become incorporated into a highly organized microbial architecture. Some microbes produce sticky glue-like substances which keep other important members of the biofilm, such as diatoms, firmly attached to the microbialite surface (Baumgartner et al. 2009). These become highly organized communities, usually with photosynthesizers at the top, and various other types of microbes are found below the surface.
What exactly does a microbialite do?
Each of the different types of bacteria, algae, and fungi perform specific functions in their environment; they are the cogs and wheels that make the microbialites extremely productive habitats (Dupraz and Visscher 2005). Through DNA sequencing technologies, scientists have uncovered the diversity of microbes contained on microbialites, which has allowed them to determine who does what in these ecosystems. Scientists have found that Cyanobacteria are the dominant photosynthesizing bacteria on marine, hypersaline, and freshwater microbialites (Couradeau et al. 2011, Foster and Green 2011, White et al. 2015). Microbialites also contain heterotrophic species, which get their energy by consuming sugars and other molecules produced by Cyanobacteria, diatoms, and algae. Yet other microbes process sulfur, nitrogen, phosphorus, and even metals (Saghai et al. 2016). Because of the high numbers of photosynthesizing microbes, microbialites play an important role in the global carbon cycle. Together, all of these processes contribute to the recycling of nutrients, an important aspect of ecosystem functioning.

Cryptic resources
Microbialites are part of the benthos, the bottom portion of marine and inland lake ecosystems, a previously overlooked component of aquatic food webs. Pelagic (open-water) organisms were previously thought to only feed on resources contained in the water column, but emerging research is pointing to the importance of benthic biofilm as a food source for these consumers (Vadeboncoeur and Power 2017). Therefore, microbialites and their productive biofilms highlight an emerging theme in ecology: unseen resources often contribute unexpectedly to the functioning of ecosystems we value and seek to protect. In fact, abundant biofilms contained on microbialites in marine and hypersaline ecosystems are likely an important food source for invertebrates (Lindsay et al. 2017, Rishworth et al. 2018).

Uncertain future
Microbialites thrive in extreme environments, but increasing human stresses threaten these rare ecosystems. Climate change, pollution, and increasing water diversions for agriculture all pose threats to the health and functioning of microbialites. The persistence of microbialites depends on a delicate balance between surrounding water properties and the composition of the resident microbial community, both of which may be altered by global change.
Of major concern is that increasing salinity and nutrient inputs will cause the important photosynthetic Cyanobacteria, the main builders of microbialites, to diminish in number, and be replaced by other microbes that may negatively impact microbialites. Lindsay et al. (2017) found that high salinity in one section of Great Salt Lake, Utah, corresponded with a reduction of the photosynthesizing Cyanobacteria and a rise in heterotrophic bacteria, a change which may hinder further microbialites growth. Similarly, in a study of Lake Clifton microbialites, Smith et al. (2010) reported that a trend of increasing salinity over the past three decades coincided with a major shift in the dominant microbialite communities. This study found a startling decline in a group of Cyanobacteria that formed the foundation of microbialite formation, and an increase in numbers of other bacteria that may not be able to maintain microbialite growth. Based on these studies, it is apparent that continued research will need to occur in a variety of marine and lake ecosystems to increase our understanding of how microbialite communities will respond to global change.
References
Baumgartner, L. K., C. Dupraz, D. H. Buckley, J. R. Spear, N. R. Pace, and P. T. Visscher. 2009. Microbial species richness and metabolic activities in hypersaline microbial mats: insight into biosignature formation through lithification. Astrobiology 9:861-874.
Bowlin, E. M., J. S. Klaus, J. S. Foster, M. S. Andres, L. Custals, and R. P. Reid. 2011. Environmental controls on microbial community cycling in modern marine stromatolites. Sedimentary Geology 263-264:45-55.
Couradeau, E., K. Benzerara, D. Moreira, E. Gérard, J. Kaźmierczak, R. Tavera, P. López-García. 2011. Prokaryotic and eukaryotic community structure in field and cultured microbialites from the alkaline Lake Alchichica (Mexico). PLoS ONE 6: e28767. doi:10.1371/journal.pone.0028767.
Dupraz, C., R. P. Reid, O. Braissant, A. W. Decho, R. S. Norman, and P. T. Visscher. 2009. Processes of carbonate precipitation in modern microbial mats. Earth-Science Reviews 96:141-162.
Dupraz, C., and P. T. Visscher. 2005. Microbial lithification in marine stromatolites and hypersaline mats. Trends in Microbiology 13:429-438.
Fernandez, A. B., M. C. Rasuk, P. T. Visscher, M. Contreras, F. Novoa, D. G. Poire, M. M. Patterson, A. Ventosa, and M. E. Farias. 2016. Microbial diversity in sediment ecosystems (evaporites domes, microbial mats, and crusts) of hypersaline Laguna Tebenquiche, Salar de Atacama, Chile. Frontiers in Microbiology 7:1284.
Foster, J. S., and S. J. Green. 2011. Microbial diversity in modern stromatolites. Life in Extreme Habitats and Astrobiology 18:383-405.
Lindsay, M. R., C. Anderson, N. Fox, G. Scofield, J. Allen, E. Anderson, L. Bueter, S. Poudel, K. Sutherland, J. H. Munson-McGee, J. D. Van Nostrand, J. Zhou, J. R. Spear, B. K. Baxter, D. R. Lageson, E. S. Boyd, Microbialite response to an anthropogenic salinity gradient in Great Salt Lake, Utah. Geobiology 2017:1-15.
Papineau, D., J. J. Walker, S. J. Mojzsis, and N. R. Pace. 2005. Composition and structure of microbial communities of Hamelin Pool in Shark Bay, Western Australia. Applied and Environmental Microbiology 71:4822-4832.
Rishworth, G. M., R. Perissinotto, M. S. Bird, and N. Pelletier. 2018. Grazer responses to variable macroalgal resource conditions facilitate habitat restructuring. Royal Society Open Science 5:171428.
Smith, M. D., S. E. Goater, E. S. Reichwaldt, B. Knott, and A. Ghadouani. 2010. Effects of recent increases in salinity and nutrient concentrations on the microbialite community of Lake Clifton (Western Australia): are the thrombolites at risk? Hydrobiologia 649:207-216.
White, R. A., I. M. Power, G. M. Dipple, G. Southam, and C. A. Suttle. 2015. Metagenomic analysis reveals that modern microbialites and polar microbial mats have similar taxonomic and functional potential. Frontiers in Microbiology 6:966 doi: 10.3389/fmicb.2015.00966
Kate received her Ph.D. in Aquatic Ecology from the University of Notre Dame and she holds a Masters in Environmental Science & Biology from SUNY Brockport. She currently teaches at a small college in Indiana and is starting out her neophyte research career in aquatic community monitoring. Outside of lab and fieldwork, she enjoys running and kickboxing.